| Inertgas-Sättigung (N2-Sättigung) | Sättigungstauchen | Blasenbildung |
| Blasenwachstum | Gastransport |
Inertgas-Sättigung
Vor dem Tauchgang
Solange sich ein Taucher im Wasser höheren Umgebungsdrucken aussetzt, sättigen sich Blut und Körpergewebe nach dem Gasgesetz von Henry entsprechend dem Umgebungsdruck mit den in der Lunge vorhandenen Gasen auf. Beim Tauchen mit Druckluft ist dabei nur die Aufsättigung mit dem Inertgas N2 von Bedeutung.
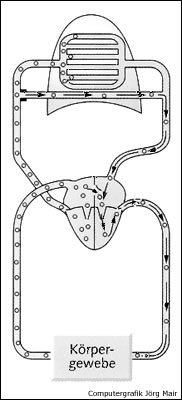
Auf Grafik 17 sind großer und kleiner Blutkreislauf mit dem äußeren Gasaustausch in der Lunge und dem inneren Gasaustausch in drei "Beispiel"-Körpergeweben dargestellt, rechtes und linkes Herz wurden der Einfachheit halber nicht eingezeichnet.
In der Lunge, im arteriellen Blut, in den drei Beispielgeweben und dem venösen Blut zeigen die grünen Balken den N2-Teildruck an. Das obere Beispielgewebe ist sehr gut durchblutet, das mittlere Gewebe hat eine mittelstarke Durchblutung und das untere Beispielgewebe eine geringe Durchblutungsrate, wie z.B. das Unterhautfettgewebe.
Aufgrund des dauernden Aufenthaltes bei einem Umgebungsdruck von 1 bar herrscht in allen Körpergeweben, im Blut und in der Lunge vor dem Tauchgang der gleiche N2-Teildruck von ca. 765 mbar (Fließgleichgewicht).
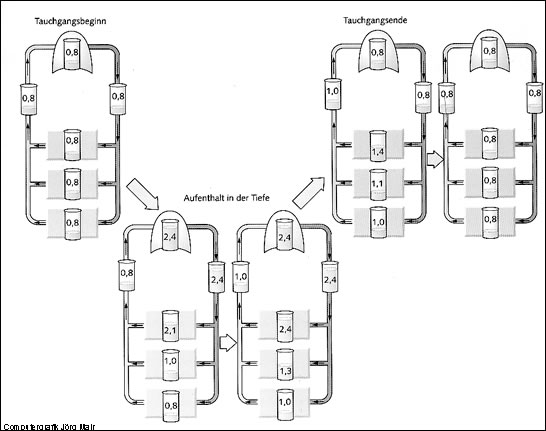
Abtauchen
Grafik 28 B zeigt die Situation kurz nach dem Erreichen einer größeren Wassertiefe. Der Teildruck des gasförmigen N2 in der Lunge stellt sich sofort entsprechend der Gesamtdruckerhöhung ein. Aufgrund der großen Austauschfläche zwischen Lunge und Blut herrscht im Blut wenige Minuten später der gleiche Teildruck des (hier allerdings gelösten) N2 wie in der Lunge.
In Abhängigkeit von der Durchblutungsrate kann das arteriell unter einem erhöhtem Teildruck angebotete N2 unterschiedlich schnell in die Gewebe diffundieren und dort zu einem Anstieg des N2-Teildrucks führen. Aufgrund der Gewebeaufsättigung liegt der N2-Teildruck im venösen Blut besonders zu Beginn eines Tauchgangs nur wenig über der N2-Sättigung unter Normalbedingungen bei 1 bar.
Aufenthalt in der Tiefe
Je länger der Aufenthalt unter erhöhtem Druck dauert, umso mehr sättigen sich die verschiedenen Gewebe mit N2 auf. Dies führt auch dazu, dass der N2-Teildruck im venösen Blut langsam ansteigt. Dauert der Aufenthalt in der Tiefe lang genug, so stellt sich wie zuvor bei 1 bar auch hier ein Fließgleichgewicht mit überall gleich hohen N2-Teildrucken ein.
Um die Sättigungsvorgänge der Körpergewebe nachvollziehen zu können, führt man Berechnungen mit "Modell"-Geweben mit jeweils unterschiedlichen Sättigungsgeschwindigkeiten durch. Eine große Rolle spielen bei diesen Berechnungen die sogenannten Sättigungshalbwertzeiten der einzelnen Gewebe.
Diese Halbwertzeiten beschreiben die Zeitspanne, die ein Gewebe nach einer Druckerhöhung benötigt, um die Hälfte der N2-Menge aufzunehmen, die dem Umgebungsdruck entspricht. Da bei großen Konzentrationsunterschieden die N2-Diffusion vom Blut ins Gewebe wesentlich schneller geschieht als bei geringen Druckunterschieden, ist ein Gewebe also nicht etwa nach zwei Sättigungshalbwertzeiten vollständig mit N2 gesättigt, sondern erst nach ca. 5 Halbwertzeiten.
Beispiel:
Beim Tauchgang auf 30 m Wassertiefe (Umgebungsdruck 4 bar) soll die N2-Aufsättigung eines Gewebes mit einer Sättigungshalbwertzeit von 20 Minuten beobachtet werden. Dieses Beispielgewebe wird so durchblutet, dass nach einem 20-minütigen Aufenthalt bei 4 bar der N2-Teildruck im Gewebe 50% des N2-Teildrucks erreicht hat, der dem Umgebungsdruck von 4 bar entspricht.
Innerhalb der nächsten 20 Minuten (Minute 21 - Minute 40) erreicht das Gewebe einen N2-Teildruck von 75 % des in 4 bar erreichbaren N2-Teildrucks, nämlich 50% der verbleibenden Differenz zwischen der akutellen Gewebesättigung und der maximal erreichbaren Gewebesättigung. In der dritten Halbwertzeit bis zur 60. Minute wird so eine relative N2-Sättigung von 87,5 % erreicht.
In der vierten Halbwertzeit bis zur 80. Minute werden 93,75 % erreicht. Nach insgesamt fünf Sättigungshalbwertzeiten werden nach 100 Minuten 96,875 % der maximal möglichen Sättigung des Gewebes mit N2 erreicht. Das heißt, dieses Gewebe ist nach 5 Sättigungshalbwertzeiten fast vollständig N2-gesättigt.
Bei der üblichen Dauer von Sporttauchgängen zwischen 20 und vielleicht 60 Minuten erreichen nur sehr gut durchblutete Gewebe mit kurzen Sättigungshalbwertzeiten eine N2-Sättigung, die meisten Gewebe werden nur zum Teil aufgesättigt. Nach heutigem Kenntnisstand gibt es im menschlichen Körper Gewebe mit Sättigungshalbwertzeiten zwischen wenigen Minuten und über 10 Stunden.
Entsättigung
Die Anpassung der N2-Sättigung der einzelnen Körpergewebe bei sinkendem Umgebungsdruck erfolgt analog der Sättigung. Auch hier reagieren gut durchblutete Gewebe mit einer schnelleren Anpassung an den aktuellen Umgebungsdruck. Schlecht durchblutete Gewebe haben noch eine gewisse Zeit lang einen N2-Teildruck, der über dem aktuellen N2-Teildruck in der Umgebungsluft liegt.
Aufgrund der Austauschfläche zwischen Lunge und Blut stellt sich im arteriellen Blut sehr schnell der aktuelle N2-Teildruck ein. Während der Durchblutung der verschiedenen Gewebe wird entsprechend dem Konzentrationsgefälle solange N2 in das Blut abgegeben, bis hier wieder ein Fließgleichgewicht auf dem ursprünglichen Niveau vor Beginn des Tauchgangs erreicht ist.
Bis zu diesem Zeitpunkt wird das überschüssige N2 über das venöse Gefäßsystem zur Lunge transportiert und dort abgeatmet. Wie die Sättigung der Gewebe so erfolgt auch die Entsättigung exponentiell, d. h. in den Geweben wird noch für ca. 5 Halbwertzeiten ein erhöhter N2-Teildruck bestehen.
Sättigungstauchen
Verweilt ein Taucher so lange in der Tiefe, bis alle seine Körpergewebe vollständig gesättigt sind, so spricht man von einem Sättigungstauchgang (engl.: Saturation-Dive). Beim Tauchen mit Pressluft wäre dieser Zeitpunkt erst nach über 2 Tagen erreicht, da die langsamsten Gewebe eine Sättigungs-Halbwertzeit von über 10 Stunden haben.
Ab Erreichen der Sättigung bleiben die erforderlichen Dekompressionszeiten für den Taucher konstant, d.h. die Relation zwischen effektiver Arbeitszeit unter Wasser und unproduktiver Dekozeit wird umso günstiger, je länger der Taucher unter Druck bleibt.
In der Offshore-Industrie (Öl-/Gasförderung etc.) führen Taucher daher oft mehrwöchige "Tauchgänge" durch. Zwischen ihren Einsätzen im Meer verbringen die Taucher dabei die Ruhezeiten in Druckkammern auf einer Bohrplattform oder an Bord eines Schiffes. Zum Arbeiten werden sie mit einer kleinen Druckkammer (engl.: "Bell") wie mit einem Fahrstuhl bis zu ihrer Arbeitstiefe gebracht.
Blasenbildung
Die Bildung von N2-Blasen im Körper ist vergleichbar mit der Bildung der CO2-Blasen in einem Glas Sekt, einem Weizenbier, CO2-haltiger Erfrischungsgetränke oder Mineralwasser. Bei all diesen Getränken löst sich bei relativ niedrigen Temperaturen eine gewisse CO2-Menge in der Flüssigkeit.
Da wärmere Flüssigkeiten weniger Gase in Lösung halten können als kühle Flüssigkeiten, werden die Flaschen mit festen Verschlüssen versehen. Das Ausgasen von CO2 aus der Flüssigkeit führt zu einer Druckerhöhung in der Flasche und bewirkt damit, dass das restliche CO2 aufgrund des erhöhten Umgebungsdrucks in Lösung gehalten wird.
Stickstoff-Blasen
Lässt man Temperaturunterschiede zunächst einmal außer Acht, so besteht gegen Ende des Tauchgangs beim Aufstieg aufgrund des sinkenden Umgebungsdrucks in den verschiedenen Körpergeweben eine relative N2-Übersättigung, abhängig von der während des Tauchgangs erfolgten Aufsättigung.
 |
Diese Übersättigung allein führt nicht zur Bildung von gasförmigem N2 in Körpergeweben oder im Blut, da zur Bildung einer Gasblase eine erhebliche Energiemenge erforderlich ist, die durch die relativ geringe N2-Sättigung nicht erbracht wird. Die Bildung von N2-Blasen ich nur möglich, wenn zur Gewebeübersättigung noch andere Faktoren hinzukommen. |
Es gibt hierzu verschiedene Überlegungen:
Scherkräfte
Zum einen kann es durch Scherkräfte im Gewebe, z.B. bei der Anspannung von Muskulatur, zu ungleichen lokalen Druckverhältnissen kommen. An den so entstehenden Orten mit einem relativen Unterdruck kann es zur Bildung von Stickstoffblasen kommen, im Gewebe oder in den Blutgefäßen.
Blasenkerne
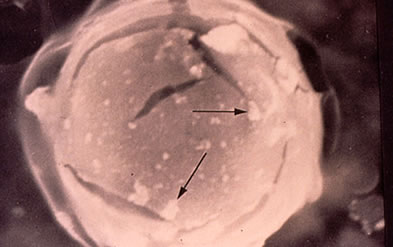
Der wahrscheinlichste Entstehungsmechanismus für N2-Blasen ist in Grafik 29 dargestellt. Zieht man den Vergleich mit dem Sektglas noch einmal heran, so kann man bei näherer Betrachung beobachten, dass die CO2-Blasen ihren Ursprung von wenigen, immer gleichen Stellen des Glasrandes haben.
 |
Es handelt sich dabei in der Regel um Unhebenheiten in der Glasoberfläche, die beim Befüllen des Glases nicht komplett mit Flüssigkeit benetzt worden sind. |
In diese wandständigen Gasblasen diffundiert das CO2 aus der übersättigten Flüssigkeit und führt solange zur Vergrößerung der wandständigen Blase, bis sich eine kleine CO2-Blase ablöst und zur Oberfläche steigt.
Die zur Zeit wahrscheinlichste Entstehungshypothese für N2-Blasen im Körper geht davon aus, dass sich in den Blutgefäßen ähnliche Unebenheiten wie im Sektglas befinden. Da sich die Gase in derartigen wandständigen Hohlräumen entsprechend dem Gesetz von Boyle-Mariotte verhalten, vergrößern oder verkleinern sie sich je nach Veränderung des Umgebungsdruckes.
Die Teildrucke der Gase entsprechen dabei immer dem aktuellen Umgebungsdruck, das heißt es gibt hier keine Über- oder Untersättigung.
Während der Dekompressionsphase eines Tauchgangs kann das in den N2-übersättigten Geweben gelöste N2 entsprechend dem Konzentrationsgefälle in eine solche wandständige Blase hinein diffundieren, da in der Blase keine N2-Übersättiung vorliegt.
Nach zunehmender Vergrößerung der wandständigen Blase durch N2-Einstrom können sich N2-Blasen ablösen und mit dem venösen Blutstrom aus dem Gewebe in Richtung rechtes Herz und Lunge transportiert werden.
Blasenwachstum
Auf ihrem Weg im venösen Blut kann eine N2-Blase durch weiteren Einstrom von N2 oder durch Zusammenschluss mehrerer kleinerer Blasen wachsen oder aber wieder schrumpfen, wobei das gesammte N2 wieder in Lösung geht. Für das Schicksal einer N2-Blase sind dabei unter anderem zwei Kräfte maßgeblich:
Die Oberflächenspannung der Gasblase und die N2-Sättigung der umgebenden Flüssigkeit. Im Inneren einer N2-Blase herrscht ein Druck, der etwas über dem Umgebungsdruck liegt, da zusätzlich zum Umgebungsdruck auch die Oberflächenspannung auf das Blaseninnere wirkt.
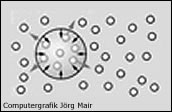 |
Entspricht der N2-Teildruck in der umgebenen Flüssigkeit dem Umgebungsdruck, so führt der relative Überdruck in der Gasblase zur Diffusion von N2 aus der Blase in die Flüssigkeit und damit zur Schrumpfung der N2-Blase. |
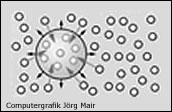 |
Erst wenn der N2-Teildruck der umgebenen Flüssigkeit höher ist als der Blaseninnendruck (Umgebungdruck plus Oberflächenspannung), kommt es durch einen weiteren N2-Einstrom in die Blase zum Wachstum. |
Gastransport
Abatmung in der Lunge
Unabhängig davon, ob das N2 im venösen Blut physikalisch gelöst ist oder in Form kleiner Gasblasen zur Lunge transportiert wird, kann das überschüssige N2 in der Regel über die Lunge abgeatmet werden. Das im Blut gelöste N2 diffundiert entsprechend dem Konzentrationsgefälle aus den Lungenkapillaren in die Lungenalveolen, wie es vom äußeren Gasaustausch des CO2 her bekannt ist.
Die Blasen des gasförmig transportierten N2 werden in den Lungenkapillaren wie in einem Filter festgehalten und führen hier zu einem lokalen Durchblutungsstop.
Das in der Blase befindliche N2 diffundiert dann wie das gelöste N2 entsprechend dem Konzentrationsgefälle in die Lungenalveolen und führt so zur Verkleinerung und schließlich zum Verschwinden der N2-Blase.
Shuntgefäße in der Lunge
 |
Wenn pro Zeiteinheit sehr viele N2-Blasen aus den Körpergeweben in die Lunge gelangen, so können bis zu 30% aller Lungenkapillaren gleichzeitig durch N2-Blasen verlegt werden, ohne dass es zu subjektiv wahrnehmbaren Symptomen kommt. Diese Situation führt allerdings durch den erhöhten Gesamtwiderstand der Lunge zu einem Blutrückstau, der zu einer Erhöhung des Blutdrucks in den Lungenarterien und zu einer Belastung des rechten Herzens führt. Überstiegt der Lungenarteriendruck ein gewisses Maß, so werden sogenannte Kurzschluss- oder Shunt-Gefäße in der Lunge geöffnet, durch die N2-reiches und blasenreiches Blut direkt in die Lungenvenen gelangt. Derartige, normalerweise nicht oder kaum durchblutete Shuntgefäße gibt es übrigens an vielen Stellen im Körper. |
Foramen ovale
In der oben beschriebenen Situation mit Erhöhung des arteriellen Drucks in der Lunge und im rechten Herz kann es bei einigen Menschen noch zu einem weiteren Effekt kommen.
Bei Menschen mit einem nicht fest verwachsenen ovalen Fenster in der Herzscheidewand kann es zum Blutübertritt aus dem rechten Vorhof in den linken Vorhof kommen, wenn der Blutdruck im rechten Vorhof den im linken Vorhof überschreitet. Normalerweise tritt dies nicht auf, da das den gesamten Körper versorgende linke Herz stets höhere Innendrücke aufweist als das rechte Herz.
Bei einer Erhöhung des Lungenwiderstandes oder bei Pressatmung mit einer Druckerhöhung im Brustraum kann jedoch der Druck im rechten Vorhof den im linken Vorhof überschreiten. Dies führt dann zum Übertritt von N2-reichem und blasenreichem Blut unter Umgehung des Lungenkreislaufs direkt in den linken Vorhof.
Arterialisierung von Blasen
Beide Möglichkeiten, geöffnete Shuntgefäße in der Lunge und Rechts-links-Shunt durch ein offenes Foramen ovale, führen so zur Einschwemmung von N2-Blasen in den Körperkreislauf und zu einer verzögerten N2-Entsättigung des Körpers, da das schon N2-vorgesättigte arterielle Blut zu einem geringeren Konzentrationsgefälle in den Geweben führt.
